
1 UVOD
Egzekutivne ili izvršne funkcije predstavljaju set povezanih, visoko kompleksnih kognitivnih veština koje su neophodne za adaptivno i cilju usmereno ponašanje jedinke. Koncept egzekutivnih funkcija se može proučavati sa različitih stanovišta, u odnosu na to koji aspekt ljudskog ponašanja je u fokusu. Koncept egzekutivnih funkcija se može proučavati iz perspektive neuro-fiziologije, neuro-nauke, psihologije i naročito razvojne psihologije i teorije ličnosti. Lista kognitivnih sposobnosti koje spadaju u niži rodni pojam u odnosu na koncept EF su: održavanje pažnje i mentalnih procesa, fleksibilnost (pažnje i mentalnih procesa), vremensku kontrolu (sekvenciranje) mentalnih procesa i ponašanja, aktivacija adekvatnih i inhibicija neadekvatnih odgovora, sposobnost autokrekcije, humor, divergenetno mišljenje, rešavanje problema, sposobnost planiranja, opštu kontrola ponašanja, kontrolu socijalnog ponašanja, kontrolu motivacije, itd.[1] Ono što objedinjuje sve egzekutivne funkcije bi se moglo opisati kao voljno, cilju usmereno, svrsishodno ponašanje.
Poslednjih decenija, raste interesovanje za proučavanje egzekutivnih funkcija, kako u okviru kognitivne psihologije, tako i u oblasti neuronauke i neurofiziologije. Izučavanje egzekutivnih funkcja kao seta međupovezanih kognitivnih procesa nastalo je kao odgovor na izjednačavanje pojmova „frontalnih funkcija“ i egzekutivnih funkcija, gde se donekle može staviti znak jednoakosti između ova dva pojma, s tim što egzekutivne funkcije prevazilazile svojim obimom funkcije vezane za frontalni korteks i njegovu ulogu u voljnom i svrsishodnom ponašanju. Razumevanje neurobioloških osnova ovih funkcija postalo je ključno za proučavanje normalnog i patološkog funkcionisanja mozga. Posebna pažnja posvećena je ulozi prefrontalnog korteksa, kao i dopaminergičkim i drugim neurotransmiterskim sistemima koji modulišu egzekutivne procese.
Cilj ovog rada je da pruži neuroanatomsku osnovu egzekutivnih funkcija, funkcionalne mehanizme, razvojne aspekte i kliničke implikacije, sa posebnim osvrtom na domapinergičke, NE, serotoninske i holinergičke mehanizme uključene u rad egzekutivnih funkcija kao i gensku ekspresiju koja je pod uticajem ovih neuronskih kola.
2 TEORIJSKI OKVIR EGZEKUTIVNIH FUNKCIJA
Literatura i istraživanja i zaključci autora koji su korišćeni kao primarni i sekundarni izvori na različite načine definišu i razumevaju koncept egzekutivnih funkcija. Ono u čemu se generalno slažu je da se koncept egzekutivnih funkcija odnosi na niz kognitivnih sposobnosti koje se nalaze visoko u hijerarhiji kognitivnih sposobnosti i njihove funkcionalizacije kojima je zajedničko da predstavljaju „dispozicije za saznajnu delatnost bilo koje vrste.“[2]
U neuropsihologiji ovaj pojam implicitno podrazumeva podelu na bazične kognitivne funkcije (u koje ubrajamo pažnju, pamćenje, percepciju, mišljenje, jezik, praksiju i slično), i egzekutivne, koje zapravo ne pripadaju ni jednom od ovih bazičnih kognitivnih domena već su njima nadređene (Jurado &Rosselli, 2007; Miyake et al., 2000). Jedan broj autora/istraživača, ipak, smatra da se domeni operisanja egzekutivnih funkcija preklapaju sa domenima pažnje, zaključivanja i rešavanja problema (Pennington & Ozonoff, 1996; Salthouse, 2005).[3]
2.1. Različiti teorijski modeli i koncepti EGZEKUTIVNE FUNKCIJE
- Lurijev model – razlikuje tri funkcionalna bloka, a treći je smešten u prefrontalni korteks i njegova funkcija je regulacija i kontrola mentalnih aktivnosti i ponašanja. PFK je integrativna jedinica, nadređena svim ostalim delovima sistema.
- Bedlijev model radne memorije – ističe komponentu centralnog izvršioca, odnosno egzekutora kompleksnih kognitivnih operacija kao što su selekcija i aktivacija kontrolnih procesa i strategija.
- Norman i Šeliz – uvode model supervizora i čine jedan od najuticajnijih modela savremene kognitivne psihologije. Celokupnu kogniciju dele na 4 sistema, a najviše mesto u hijerarhiji ima četvrti i najviši koji nazivaju SAS – supervisory attentional system koji je fleksibilan i visoko adaptibilan u novim situacijama. SAS je prema njihovom konceptu sukcija frontalnih režnjeva.
- Štrus i Benson – razvili su tripartitni model. Egzekutivne funkcije posreduju u razvijanju i održavanju veza koje postoje između frontalnih regiona, limbičkih i posteriornih regiona.
- Lezak Mjuriel – njen model je jedan od najprihvaćenijih modela egzekutivnih funkcija. Sama ga je neznatno promenila i dopunila nakon 1995. godine, razlikujući 4 sržne komponente egzekutivnih funkcija: htenje, planiranje, svrhovitu aktivnost i efikasnost postignuća.
- Barkli – uvodi model egzekutivne self-regulativne funkcije. Njegov model egzekutivne funkcije razlikuje četiri primarna domena EF: radnu memoriju, samo-regulaciju, internalizovani govor i rekonstrukciju ili bihejvioralnu sintezu.
- Dankan – teorija zanemarivanja cilja. Srž njegovog teorijskog modela je značaj kontrole aktivacije i inhibicije ponašanja koje nam olakšavaju ili otežavaju obavljanje zadataka.
- Mijake i saradnici – izdvojili su najmanje tri egzekutivne funkcije koje se ne mogu dalje svesti jedna na drugu a nisu nezavisne već čine jednu funkcionalnu i organizacionu celinu ili sistem. Oni razlikuju: premeštanje pažnje (set-shifting), inhibiciju automatskih odgovora i ažururanje radne memorije ulaznim i izlaznim podacima. Pored ove tri osnovne, razlikuju i verbalnu fluentnost.
Ovi modeli pružaju osnovu za razumevanje kako se egzekutivne funkcije razvijaju, kako se manifestuju u ponašanju i kako mogu biti narušene u različitim kliničkim stanjima, kao što su poremećaji pažnje, autizam, šizofrenija, depresija, bipolarni poremećaj. Možemo ih podeliti na dva osnovna modela:
- Hijerarhijski modeli prefrontalnog korteksa koji pretpostavljaju da različite regije PFK obavljaju različite nivoe kontrole ponašanja jedinke.
- Integrativni modeli koji povezuju egzekutivne funkcije sa emocionalnom regulacijom, motivacijom i kognicijom.
Ovi modeli pružaju osnovu za razumevanje kako se egzekutivne funkcije razvijaju, kako se manifestuju u ponašanju i kako mogu biti narušene u različitim kliničkim stanjima.
2.2. Hladne i vruće egzekutivne funkcije
U savremenoj literaturi pravi se razlika između tzv. „hladnih“ i „vrućih“ egzekutivnih funkcija:
- Hladne EF odnose se na apstraktne, logičke i kognitivno neutralne zadatke (npr. rešavanje problema, planiranje).
- Vruće EF uključuju emocionalno obojene situacije koje zahtevaju donošenje odluka pod uticajem nagrade, kazne ili afekta (npr. odlaganje zadovoljenja, moralno rasuđivanje).
Ova podela ima važne implikacije za razumevanje ponašanja u realnim životnim situacijama, kao i za dijagnostiku i tretman neuropsihijatrijskih poremećaja. U popualrnoj psihologiji i razvoju pojednica ima svoje mesto kada se dizajniraju treninzi za meke i tvrde veštine.
2.3. Vremenska skala sazrevanja prefrontalnog korteksa i razvoj egzekutivnih funkcija[4]
Razvojne studije daju ubedljive podatke da različite komponente egzekutivnih funkcija sazrevaju u različitim periodima, počevši od ranog detinjstva sve do (bar) sredine adolescencije.[5]
Prefrontalni korteks je deo mozga koji se najsporije razvija. On dostiže punu zrelost tek oko 25. godine života. Sporo sazrevanje PFK koristi se kao argument za objašnjenje zašto se egzekutivne funkcije razvijaju postepeno u ciklusima i talasima. Prate razvoj i rast jedinke, mada se ne može uvek staviti znak jednakosti između fiziološkog odrastanja i sazrevanja egzekutivnih funkcija.
Ključni neurofiziološki procesi koji se odvijaju:
- Sinaptogeneza (porast broja sinapsi) u ranom detinjstvu;
- Sinaptičko orezivanje (pruning) tokom adolescencije čime se kognitivne mreže specijalizuju, a važne su za razvoj egzekutivnih fukncija, i;
- Mijelinizacija koja je važna za brzu obradu informacija i regulaciju impulsa.
Ovi procesi direktno utiču na to kada i kako se pojedine komponente egzekutivnih funkcija pojavljuju.
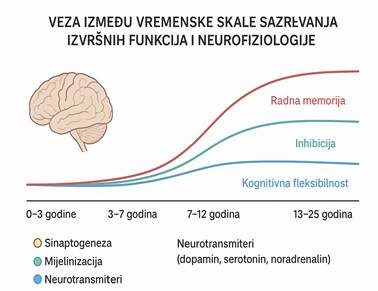
Slika 1 (generisana pomoću AI na osnovu podataka iz Vicki Anderson, Rani Jacobs, Peter J Anderson, Executive functions and the frontal lobes: A lifespan perspective, Taylor and Frances, 2008)
2.3.1. Ontogenetski razvoj egzekutivnih funkcija
Razvoj egzekutivnih funkcija je usko povezan sa maturacijom prefrontalnog korteksa, koji se razvija najsporije od svih moždanih regija:
- Rano detinjstvo (3–6 godina): Početak razvoja inhibicije i radne memorije. Deca počinju da kontrolišu impulse i planiraju jednostavne zadatke.
- Srednje detinjstvo (7–12 godina): Značajan napredak u kognitivnoj fleksibilnosti, organizaciji i samoregulaciji. Školski zahtevi dodatno stimulišu razvoj egzekutivnih funkcija.
- Adolescencija: (13-19): Nastavak razvoja apstraktnog mišljenja, donošenja odluka i emocionalne regulacije. Povećana aktivacija ventromedijalnog PFK-a u emocionalnim zadacima.
- Rana odrasla dob: (20-29): Dostiže se vrhunac efikasnosti egzekutivnih funkcija, uz stabilizaciju funkcionalnih mreža i neurotransmiterske ravnoteže.
- Starenje: U starijem dobu dolazi do postepenog opadanja egzekutivnih funkcija, posebno u domenima brzine obrade, radne memorije i fleksibilnosti. Međutim, iskustvo i kompenzatorni mehanizmi mogu ublažiti efekte starenja.
3 Neuroanatomija egzekutivnih funkcija
Egzekutivne funkcije su prvenstveno povezane sa aktivnošću prefrontalnog korteksa (PFK), koji predstavlja najrazvijeniji deo ljudskog mozga i ključnu strukturu za regulaciju složenih kognitivnih procesa. Neuroanatomske studije, funkcionalna neuroimaging istraživanja i neuropsihološki testovi dosledno ukazuju na to da različite regije PFK-a imaju specifičnu ulogu u ispoljavanju egzekutivnih funkcija. Analiza Pijažeovih zadataka operativnog mišljenja koje prate ontogenezu PFK i testova za specifične egzekutivne funkcije pokazuje povezanost između postignuća na ovim zadacima, odnosno, operativno mišljenje zavisi od razvijenosti egzekutivnih funkcija, t.j. od zrelosti PFK i njegovih funkcija. Činjenica da može da se uspostavi veza između ova dva koncepta ne dokazuje uzročnost. Relevantnost sazrevanja prefrontalnih struktura za funkcije kao što su radna memorija ili inhibitorna kontrola motornih akcija dokazale su Dajmond i Goldman-Rakić serijom eksperimenata u kojima su poredile postignuće na Pijažeovim probama ranih saznajnih procesa kod dece i viših primata sa eksperimentalnim prefrontalnim lezijama (Diamond, 1985, 1991; Diamond i Goldman-Rakic, 1989; Goldman-Rakic 1987b).[6]
3.1. Prefrontalni korteks
Prefrontalni korteks se funkcionalno deli na više regija, od kojih svaka ima specifičnu ulogu u egzekutivnim funkcijama:
- Dorzolateralni prefrontalni korteks (dLPFK): Uključen u radnu memoriju, planiranje, rešavanje problema i kognitivnu fleksibilnost. Aktivacija DLPFK-a se beleži tokom zadataka koji zahtevaju manipulaciju informacijama i donošenje odluka bez emocionalnog sadržaja. Zauzima većinu spoljne strane prefrontalnih oblasti, uključujući uglavnom lateralne strane BA 9-12, celokupne BA 45, 46 i gornje oblasti aree 47 (BA 12 i 45 se preciznije označavaju i kao ventrolateralne)
- Ventromedijalni prefrontalni korteks (vMPFK)/ Orbitofrontalni korteks (OFK):
Povezan sa donošenjem odluka u emocionalno obojenim kontekstima, evaluacijom nagrade i kazne, kao i socijalnim rasuđivanjem. Oštećenja ove regije dovode do impulsivnosti i poremećaja moralnog rasuđivanja. Uključen je u inhibiciju neadekvatnih odgovora, evaluaciju nagrade i adaptaciju ponašanja na osnovu povratne informacije. OFK igra ključnu ulogu u „vrućim“ egzekutivnim funkcijama. Orbitofrontalni korteks predstavlja mrežu koja učestvuje u bihejvioralnoj kontroli vezanoj za socijalne situacije, ali i u onim komponentama ponašanja koje je Lermit nazvao “nezavisnošću od sredine”; a dorzolateralni korteks predstavlja mrežu koja podržava egzekutivne funkcije i programiranje (mentalnih i motornih) akcija individue (Damasio, 1985; Grattan i Eslinger, 1991; McPherson i Cummings, 2002).[7] - Anteriorni cingulatni korteks (ACC) anteriorne oblasti imaju značajniju ulogu u jezičkoj produkciji, a posteriorne u razumevanju.[8] ACC je uključen u detekciju grešaka, monitoring konflikta i regulaciju pažnje. Njegova aktivacija se beleži u zadacima koji zahtevaju izbor između konkurentnih odgovora, kao što su Stroop test[9] i Go/No-Go paradigme. ACC funkcioniše kao „alarmni sistem“ koji signalizira potrebu za pojačanom kontrolom.
3.2. Bazalne ganglije
Bazalne ganglije, naročito strijatum, učestvuju u selekciji odgovora, motoričkoj inhibiciji i učenju zasnovanom na nagradi. One formiraju funkcionalne petlje sa PFK-om koje omogućavaju integraciju motivacionih i kognitivnih informacija. Sve petlje uključuju iste bazične anatomske strukture (bazalne ganglije. Prefrontalne oblasti su jedini deo kore ( osim premotorne) koje, preko talamusa, primaju input iz bazalnih ganglija. Pošto projekcije kroz bazalne ganglije imaju ulogu u više kognitivnih procesa, uključujući sekvencijalno planiranje i monitoring ponašanja, smatra se da one bitno doprinose egzekutivnoj funkciji prefrontalne kore… ali su projekcije svake od njih uglavnom razdvojene od ostalih. Svaka od petlji ima bar dva paralelna puta projekcije (direktni i indirektni), a sve strukture obuhvaćene petljom primaju informacije i iz drugih cerebralnih oblasti, funkcionalno povezenih sa specifičnom petljom ka kojoj se projektuju (Cummings, 1993).[10]
3.3. Cerebelum
Iako tradicionalno povezan sa motoričkom koordinacijom, cerebelum je sve više prepoznat kao važan za vremensko usklađivanje kognitivnih procesa, automatizaciju ponašanja i predviđanje ishoda ponašanja. Njegove veze sa PFK-om doprinose efikasnosti egzekutivne kontrole. Cerebelarne strukture imaju višestruku ulogu za kognitivno funkcionisanje: dok je klasično smatran prvenstveno centrom za senzornu regulaciju voljnih pokreta, danas se specifikuje njegova posebna uloga u motornom i prostornom učenju, izvodjenju serijskih pokreta, detekciji grešaka kod nemotornog učenja kao i, globalnije, u modulaciji viših kognitivnih funkcija.[11]
4. NEUROTRANSMITERI
Neurotransmiteri su molekuli koji nastaju u presinaptičkim neuronima, u vezikulama u nervnoj ćeliji, a procesom egzocitoze se oslabađaju u sinaptičkoj pukotini, vezuju se za receptore koji se nalaze u membrani postsinaptičke ćelije i dalje menjaju aktivnost postinaptičke ćelije na specifičan način. Mehanizmi koji utiču na prenos hemijskog signala sa pre- na postsinaptičku ćeliju su različiti: akcionim potencijalom, jonskim i metabotropnim mehanizmima, ili posredno preko neuromodulatora.
4.1. Uloga neurotransmitera
Neurotransmiteri igraju ključnu ulogu u regulaciji egzekutivnih funkcija, posebno u prefrontalnom korteksu:
- Dopamin: zauzima centralno mesto za radnu memoriju, motivaciju i fleksibilnost u donošenju odluka. Dopaminergička aktivnost u dorzolateralnom PFK-u povezana je sa efikasnošću u zadacima koji zahtevaju manipulaciju informacijama. Disbalans dopamina se povezuje sa poremećajima kao što su ADHD i šizofrenija.
- Noradrenalin(norepinefrin): Uključen u regulaciju pažnje, budnosti i inhibicije impulsa. Noradrenergički sistem moduliše odgovor na stres i omogućava adaptivnu kontrolu ponašanja u zahtevnim situacijama.
- Serotonin: Ima važnu ulogu u emocionalnoj regulaciji, impulsivnosti i donošenju odluka u kontekstu nagrade i kazne. Serotoninski disbalans se često povezuje sa depresijom i poremećajima raspoloženja.
- GABA (gama-aminobuterna kiselina): Glavni inhibitorni neurotransmiter u centralnom nervnom sistemu, smanjuje anksioznost i smiruje aktivnost neurona.
- Acetilholin: Značajan za učenje, pamćenje i mišićnu kontrakciju.
4.2.1. Dinamika dopmaninergičkih sistema
Dopamin učestvuje u mnogim procesima pored egzekutivnih funkcija, poput kodiranja nagrada i zavisnosti od droga (Everitt et al. 1999; Grace 1995; Koob 1992). Medijalni prefrontalni korteks (mPFK) i orbitofrontalni korteks (OFK) dobijaju dopaminergički ulaz iz ventralnog tegmentalnog područja (VTA) i ispoljavaju više podtipova dopaminskih receptora: D1, D2, D3 i D4 (Gaspar et al. 1995; Wedzony et al. 2000). Loše izvođenje zadataka set-shifting-a i pažnje javlja se kada je dopaminska aktivnost u mPFK niska, dok porast dopamina poboljšava ove procese. Farmakološka deplecija dopamina narušava set-shifting i pažnju u zadacima kod primata i glodara (Crofts et al. 2001; Robbins i Roberts 2007). Receptori D1 i D2 učestvuju u set-shifting procesima, što potvrđuje činjenica da blokatori ovih receptora u mPFK narušavaju fleksibilnost (Floresco et al. 2006; Ragozzino 2007). Suprotno tome, antagonizam D4 receptora u mPFK poboljšava set-shifting, dok agonizam D4 receptora narušava fleksibilnost (Floresco et al. 2006).[12]
4.2.2 Dinamika noradrenalinskih sistema
Locus coeruleus (LC) predstavlja glavni izvor noradrenalinskih projekcija ka mPFK i OFK. Noradrenalinski receptori — α1, α2 i β — locirani su presinaptički na noradrenergičkim aksonima, dok su α2 receptori prisutni i postsinaptički (U’Prichard et al. 1979). Od tri podtipa α2 receptora (α2A, α2B, α2C), α2A je najzastupljeniji u PFK. On se izražava na noradrenergičkim terminalima, ali i na piramidalnim neuronima (Aoki et al. 1998). NE utiče na sva četiri procesa egzekutivnog funkcionisanja jer reguliše opšti nivo pobuđenosti i podešava optimalnu aktivnost PFK u skladu sa spoljnim uslovima.[13]
4.2.3. Dinamika serotoninskih sistema
Dorzalni raphe (DR) predstavlja primarni izvor serotoninskih (5-HT) projekcija ka mPFK-u i OFK-u. Od sedam podfamilija 5-HT receptora, prefrontalni korteks izražava samo podfamilije 5-HT1, 5-HT2 i 5-HT3, a nivo njihove ekspresije varira u zavisnosti od kortikalnog sloja i tipa neurona. I mPFK i OFK šalju eferentne projekcije ka dorzalnom rapheu, što znači da oba regiona mogu direktno modulirati serotoninsku aktivnost (Enge i sar., 2011). Prema Cools i sar. (2008), serotoninska aktivnost u OFK-u je čvrsto povezana sa stepenom inhibicije odgovora i performansama u zadacima reverzibilnog učenja.[14]
4.2.4. Dinamika holinergičkog sistema
mPFK i OFC primaju difuzni holinergički input iz nucleus basalis Meynerti u bazalnom prozencefalonu. Postoje dve klase holinergičkih receptora koji utiču na egzekutivne funkcije:
- Muskarinski (mAChR) – metabotropni, spori
- Nikotinski (nAChR) – jonski, brzi
4.2.4.1. Muskarinski receptori
Njihova uloga u izvršnim funkcijama je manje jasna.
4.2.4.2. Nikotinski receptori (nAChR)
Oni igraju ključnu ulogu u: pažnji, set-shifting-u, reverzibilnom učenju i inhibiciji odgovora.[15]
Holinergički sistem, prvenstveno delujući preko presinaptičkih nikotinskih receptora, može koordinisati sve četiri izvršne funkcije posredovane mPFK-om i OFK-om:
- U mPFK-u: oslobađanje DA i NE utiču na set-shifting i pažnju;
- U OFK-u: oslobađanje NE koji utiče na inhibiciju odgovora i reverzibilno učenje, i;
- Indirektno: aktivacija nAChRs u VTA, LC i DR menja dopaminergičku, noradrenergičku i serotonergičku aktivnost
5 GENSKA EKSRESIJA i EGZEKUTVE FUNKCIJE
Genetski polimorfizam, odnosno male razlike u eksresiji gena među jedinkama, a posebno onih koji utiču na dopaminergički sistem (npr. COMT – enzim koji razgrađuje domapin u PFK-u), povezani su sa razlikama u efikasnosti egzekutivnih funkcija kod različitih jedinki. Takođe, individualni profil ekzekutivnih sposobnosti može da zavisi ne samo od gena nego i od kombinacije gena i sredinskih faktora. Genski polimorfizam je mala genetska razlika među ljudima koja objašnjava zašto se razlikujemo u osobinama, reakcijama na lekove, metabolizmu, neurohemiji, riziku za različite poremećaje.
Primarni izvor korišćen u ovom radu objašnjava ulogu domapina i modulaciju aktivnosti dopamina u odnosu na genski polimorfizam.
5.1. Dopamin
Za razliku od drugih hemikalija u mozgu, dopamin funkcioniše preko G-proteinsih veza u postsinaptičkoj ćeliji. Ovi GPRC – G protein coupled receptors mogu da utiču na gensku ekspesiju. Na primer, neurotransmiter se vezuje za receptor. U drugom koraku G-protein aktivira adenil ciklazu i na taj način nastaje cAMP. U sledećem koraku cAMP aktiviše PKA koja fosforiliše CREB. CREB dalje moduliše transkripciju gena. Geni dalje moduliraju proteine koji utiču na transmisiju signala. Proizvod ove aktivnosti je nastanak novo sintetisanog proteina, odnosno jonski kanal ili enzim modulator.[16]
5.1.1.Polimorfizam COMT Val158Met (rs4680) utiče na razgradnju dopamina u prefrontalnom korteksu, pri čemu Val alel povećava, a Met alel snižava aktivnost enzima, što rezultira nižim, odnosno višim nivoima dopamina. Istraživanja na zdravim odraslima pokazuju da nosioci Met/Met genotipa imaju bolju održanu pažnju i efikasniju neuralnu aktivaciju u ACC-u, dok Val/Val nosioci pokazuju lošije performanse; međutim, kod adolescenata se optimalne performanse beleže kod Val/Met genotipa, verovatno zbog razvojne osetljivosti PFK-a i nižih dopaminskih zahteva, dok kod starijih osoba Val/Val genotip može biti povezan sa boljom selektivnom pažnjom, što ukazuje na promene dopaminske osetljivosti tokom životnog ciklusa. Kada je reč o kognitivnoj fleksibilnosti, zdravi Met/Met pojedinci postižu bolje rezultate na WCST-u u poređenju sa ostalim genotipovima, dok kod osoba sa šizofrenijom nema razlike među genotipovima; suprotno tome, kod bipolarne manije Val/Val genotip je povezan s boljom fleksibilnošću jer visoka COMT aktivnost može ublažiti povišene dopaminske nivoe karakteristične za maniju.
5.1.2. Polimorfizmi DA receptora i izvršne funkcije
Polimorfizmi dopaminskih receptora DRD2 i DRD4 povezani su s varijacijama u egzekutivnim funkcijama kroz modulaciju fronto-strijatalnih i prefrontalnih dopaminskih mehanizama. Kod DRD2 gena, prisustvo A1+/T alela (rs1800497, DRD2/ANKK1 Taq Ia) dovodi do smanjenja gustine DRD2 receptora u strijatumu, što je povezano s boljom kognitivnom fleksibilnošću i smanjenom funkcionalnom povezanošću u dorzalnim fronto-strijatalnim krugovima, dok osobe bez ovog alela imaju gušću DRD2 receptoriku i slabije rezultate u zadacima promene strategije. Kod DRD4 gena, varijanta sa sedam ponavljanja u egzonu 3 smanjuje osetljivost receptora na dopamin, ali je uprkos tome — ili upravo zbog toga — povezana s poboljšanom kognitivnom fleksibilnošću i boljom inhibicijom odgovora, što je potvrđeno boljim performansama nosilaca ove varijante u go/no-go zadacima.
5.2. Polimorfizmi u genima povezanim sa noradrenergičkom neurotransmisijom
U noradrenergičkim neuronima, gen DβH kodira enzim dopamin β-hidroksilazu, koja katalizuje pretvaranje dopamina u noradrenalin (NE) i igra ključnu ulogu u održavanju ravnoteže između ova dva neurotransmitera u korteksu. Polimorfizmi u DβH genu povezani su sa varijacijama u aktivnosti ovog enzima. T alel, koji smanjuje transkripciju gena i usporava pretvaranje dopamina u noradrenalin, povezan je sa slabijim rezultatima u zadacima održane pažnje (Greene et al. 2009). Ovi podaci su u skladu sa nalazima da smanjenje noradrenalina u PFK-u dovodi do smanjene pažnje.
5.3. Polimorfizmi u genima povezanim sa serotonergičkom neurotransmisijom
Polimorfizam 1438 u genu za 5-HT2A receptor utiče na ekspresiju funkcionalnih 5-HT2A receptora u korteksu, a nosioci A-1438A alela pokazuju slabiju inhibiciju odgovora na „go/no go“ zadacima u odnosu na G-1438G genotip, verovatno zbog smanjene ekspresije ovih ekscitatornih receptora na piramidalnim neuronima — što je u skladu sa nalazima da aktivacija 5-HT2A receptora u orbitofrontalnom korteksu poboljšava inhibiciju odgovora. Pored toga, polimorfizmi u transporteru serotonina (5HTTLPR i varijacije u 3’ nekodirajućem regionu) utiču na kognitivnu fleksibilnost merenu obrnutim učenjem kod rezus majmuna: jedinke sa dve kopije kratkog alela 5HTTLPR pokazuju znatno slabije rezultate, dok majmuni sa T:G:T haplotipom, povezanim sa nižom ekspresijom transportera i verovatno smanjenim ponovnim unosom serotonina, prave manje grešaka u obrnutom učenju, što se uklapa u nalaze da viši nivo serotoninske aktivnosti u OFK poboljšava fleksibilnost u učenju.
5.4. Polimorfizam u genu povezanom sa holinergičkom neurotransmisijom
Gen CHRNA4, koji kodira α4 podjedinicu α4β2 nikotinskih acetilholinskih receptora, sadrži SNP rs1044396, koji podrazumeva zamenu nukleotida C → T. Ova varijanta može uticati na razlike u pažnji, a pregled literature Greenwood i saradnika (2012) pokazuje da je T alel povezan s boljom pažnjom i pojačanom kortikalnom aktivnošću, što je u skladu sa širim nalazima da nikotin, putem ovih receptora, poboljšava pažnju.
6 METODE ISTRAŽIVANJA EGZEKUTIVNIH FUNKCIJA U NEURONAUCI
- fMRI, MEG, EEG – funkcionalna dinamika
- PET – neurotransmiteri
- Optogenetika, hemogenetika (za životinje)
- TMS / tDCS – modulacija PFC funkcija
- Testovi: Stroop, WCST, Go/No-Go, Hanojska kula, Londonska kula, Kula Toronta
7 POREMEĆAJI SA IZRAŽENIM DEFICITOM EGZEKUTIVNIH FUNKCIJA
- ADHD (poremećaj pažnje sa hiperaktivnošću): Karakteriše se deficitom inhibicije, radne memorije i održavanja pažnje. Deca sa ADHD-om često imaju teškoće u organizaciji zadataka, kontroli impulsa i emocionalnoj regulaciji.
- Šizofrenija: U ovom poremećaju dolazi do značajnog oštećenja radne memorije, planiranja i fleksibilnosti mišljenja. Disfunkcija dorsolateralnog PFC-a povezana je sa negativnim simptomima i smanjenom funkcionalnošću.
- Depresija: Osobe sa depresijom pokazuju smanjenu motivaciju, rigidnost u mišljenju i teškoće u donošenju odluka. EGZEKUTIVNE FUNKCIJE disfunkcija se često održava i nakon povlačenja afektivnih simptoma.
- Bipolarni poremećaj: Tokom maničnih epizoda dolazi do impulzivnosti i smanjene inhibicije, dok depresivne faze karakteriše usporeno procesuiranje i slabija fleksibilnost.
- Neurodegenerativni poremećaji (npr. Alchajmerova i Parkinsonova bolest): U ranim fazama Alchajmerove bolesti javljaju se teškoće u planiranju i rešavanju problema, dok Parkinsonova bolest često uključuje deficit inhibicije i motoričke kontrole, usled disfunkcije bazalnih ganglija.
8 ZAKLJUČAK
Egzekutivne funkcije su skup viših kognitivnih procesa koji omogućavaju planiranje, inhibiciju i aktivaciju, fleksibilnost mišljenja, donošenje odluka i shodno tome cilju usmereno ponašanje. Egzekutivne funkcije obuhvataju složene kognitivne procese koji su nam neophpdni za svakodnevno funkcionisanje, učenje i pamćenje, primenu naučenog u novim okolnostima, kretanje, adaptaciju na stalno promenljivo spoljašnje okruženje, neurološko i mentalno zdravlje.
Nakon pročitane primarne i sekundarne literature moj lični zaključak je da su EGZEKUTIVNE FUNKCIJE složen koncept internih mentalnih kapaciteta jedinke da usmerava, prilagođava i samo-reguliše svoje ponašanje i obrasce ponašanja kako bi postigla određeni cilj. Ne mislim na kvalitet ili sadržaj ciljeva već kapacitet da se pokrene u smeru ostvarivanja svojih ciljeva. Odnosno, što sam dublje ulazila u temu egzekutinih funkcija, neuronauke, neuroanatomije, neurofiziologije i neurohemije, ovaj koncept je delovao kao da izmiče sveobuhvatnom razumevanju.
Literatura:
- Milkica Nešić, Ilustrovana fiziologija nervnog sistema, Filozofski fakultet Niš, 2013.
- Žarko Trebjašanin, Žarko Trebješanin, Rečnik psihologije, Stubovi kulture, Beograd, 2001.
- Nadežda S. Krstić, Uvod u neuropsihologiju, https://www.fasper.bg.ac.rs/do2016/nastavnici/Krstic_Nadezda/krstic.html
- Željka Nikolašević, Nasledni i sredinski činiioci kognitivnih sposobnosti, Doktorska disertacija, Univerzitet u Novom Sadu, Filozofski fakultet, 2016.
- Vicki Anderson, Rani Jacobs, Peter J Anderson, Executive functions and the frontal lobes: A lifespan perspective, Taylor and Frances, 2008.
- Nadežda Krstić, Olivera Aleksić, Predrag Vidović, Mirjana Gojković, Neurokognitivni razvoj kod dece mlađeg školskog uzrasta (I): egzekutivne funkcije, konstruktovne sposobnosti i pamćenje, Psihijat.dan./2002/34/3-4/305-331/
- Nadežda Krstić, Razvojna neuropsihologija, Univerzitet u Beogradu Fakultet za specijalnu edukaciju i rehabilitaciju, Beograd, 2008.
- Sheree F Logue, Ph.D, Thomas J Gould, Ph.D The Neural and Genetic Basis of Executive Function: Attention, Cognitive Flexibility, and Response Inhibition, Pharmacol Biochem Behav. 2014 August
- Marko Ž. Živanović, Efekti transkranijalne neuromodulacije frontoparijetalne funkcionalne mreže na više kognitivne funkcije, doktorska disertacija, Beograd, 2019.
[1] Nadežda S. Krstić, Uvod u neuropsihologiju, https://www.fasper.bg.ac.rs/do2016/nastavnici/Krstic_Nadezda/krstic.html, str. 143.
[2] Žarko Trebješanin, Rečnik psihologije, Stubovi kulture, Beograd, 2001. str. 220.
[3] Željka Nikolašević, Nasledni i sredinski činiioci kognitivnih sposobnosti, Doktorska disertacija, Univerzitet u Novom Sadu, Filozofski fakultet, 2016, str. 29.
[4] Za više informacija o vremenskoj skali maturacije PFK i veze razvoja mozga i egzekutivnih funkcija, pogledati: Vicki Anderson, Rani Jacobs, Peter J Anderson, Executive functions and the frontal lobes: A lifespan perspective, Taylor and Frances, 2008.
[5] Nadežda Krstić, Olivera Aleksić, Predrag Vidović, Mirjana Gojković, Neurokognitivni razvoj kod dece mlađeg školskog uzrasta (I): egzekutivne funkcije, konstruktovne sposobnosti i pamćenje, Psihijat.dan./2002/34/3-4/305-331/, str. 321
[6] Nadežda Krstić, Razvojna neuropsihologija, Univerzitet u Beogradu Fakultet za specijalnu edukaciju i rehabilitaciju, Beograd, 2008, str. 42.
[7] Nadežda Krstić, Uvod u neuropsihologiju, str. 152.
[8] Nadežda Krstić, Razvojna neuropsihologija, Univerzitet u Beogradu Fakultet za specijalnu edukaciju i rehabilitaciju, Beograd, 2008, str.128
[9] Pored Stroop testa, razvijeno je nekoliko tipova testova kojima se mere EF kod dece i odraslih: Trial Making Test (TMT), WCST – Viskonsin test sortitanja karata, Test verbalne fluentnosti, Ternetov test fluentnosti reči, COWAT – Test kontrolisanog usmenog asociranja reči, Hanojska kula, Kula Toronta.
[10] Nadežda Krstić, Uvod u neuropsihologiju, str. 151
[11] Nadežda Krstić, Uvod u neuropsihologiju, str. 112
[12] Prema Sheree F Logue, Ph.D, Thomas J Gould, Ph.D The Neural and Genetic Basis of Executive Function: Attention, Cognitive Flexibility, and Response Inhibition, Pharmacol Biochem Behav. 2014 August, str. 4-5.
[13] Prema Sheree F Logue, Ph.D, Thomas J Gould, Ph.D The Neural and Genetic Basis of Executive Function: Attention, Cognitive Flexibility, and Response Inhibition, Pharmacol Biochem Behav. 2014 August, str. 6-7.
[14] Prema Sheree F Logue, Ph.D, Thomas J Gould, Ph.D The Neural and Genetic Basis of Executive Function: Attention, Cognitive Flexibility, and Response Inhibition, Pharmacol Biochem Behav. 2014 August, str. 7-8.
[15] Prema Sheree F Logue, Ph.D, Thomas J Gould, Ph.D The Neural and Genetic Basis of Executive Function: Attention, Cognitive Flexibility, and Response Inhibition, Pharmacol Biochem Behav. 2014 August, str. 8-9.
[16] Milkica Nešić, Ilustrovana fiziologija nervnog sistema, Filozofski fakultet, Niš, 2013, str. 121.
